
Морфоструктура органов иммунитета промышленной птицыУДК: 619:[612.118]:636.5 Михайлов Е.В., Шабунин Б.В., Степанов Е.М. Федеральное государственное бюджетное Введение. Птицеводство - одно из наиболее устойчивых и динамичных отраслей промышленного комплекса Российской Федерации. Исходя из международных организаций к 2025 году, интегрирование производства мяса птицы будет занимать первое место в мире. Ежегодный рост производства продукции птицеводства в нашей стране сохраняется на уровне 17-19% с перспективой повышения темпов производительности. Это большой биологический потенциал, который следует рассматривать как совокупный биологический организм, требующий постоянного внимания к своему здоровью, а также поддержке и преумножению развития генетического потенциала данного организма. Отсюда следует однозначный вывод - вопрос сохранности поголовья и предотвращения болезней птиц никогда не потеряет своей актуальности.
Интенсивное развитие птицеводства обусловлено техническим переоснащением отрасли с использованием ресурсосберегающих технологий, введением новых высокопродуктивных кроссов и направлений. В период адаптации к интенсивным технологиям выращивания птица испытывает многочисленные воздействия факторов внешней среды, и чтобы обеспечить согласованное функционирование всех физиологических систем, вынуждена активизировать все защитные силы организма. Как правило, в обеспечении иммунного ответа главенствующую роль играют основополагающие органы иммунного ответа. Иммунная система объединяет органы и ткани, которые осуществляют защитные реакции организма, обеспечивая тем самым резистентность макроорганизма. В соответствии со своей функцией они делятся на центральные и периферические. У птиц к центральным органам относятся клоакальная (Фабрициева) сумка (Bursa fabricii), костный мозг, тимус (зобная, вилочковая железа), а к периферическим - железа третьего века (Гардерова), лимфоидный дивертикул (Меккеля), лимфоидные бляшки слепых кишок, селезенка (депо крови) и лимфоидные узелки, расположенные по ходу лимфатических сосудов. Ввиду повышенной мобильности иммунной системы, её активному ответу на чужеродные агенты окружающей среды, специалисты разрабатывают комплексы мер, направленных на коррекцию данной системы. Для детального понимания сущности явлений, протекающих в иммунной системе птиц, следует детально разобраться в её морфологии, физиологии и функциональном иммуностазе, поскольку нор-мергический ответ, как иммунной системы, так и организма в целом, способствует повышению продуктивности птицы, качества мяса и яйца, а также способствует преумножению генетического потенциала заведомо продуктивной птицы [7].
Материалы и методы исследований. Для изучения органов кроветворения и иммуногенеза были использованы цитологический и гистологический методы. Материалом для цитологического метода служили мазки-отпечатки, полученные с поверхностей разрезов органов. Окраску мазков проводили по методике Май Грюнвальд-Гимза, после окраски препараты были заключены в полистирол под покровное стекло для сохранности. Материалом для гистологического метода являлись кусочки органов. Фиксация происходила в 10%-нейтральном забуференном формалине. Затем материал обезвоживался в спиртах возрастающей крепости, после чего заливался в гистологический парафин «Histomix». С помощью микротома из материала изготавливались срезы органов толщиной 3-4 мм, которые расправлялись в водяной бане и помещались на высокоадгезивные стекла. Для окраски препаратов использовали обзорную окраску гематоксилином-эози-ном и трехцветную окраску Азур-2 Эозином [4].
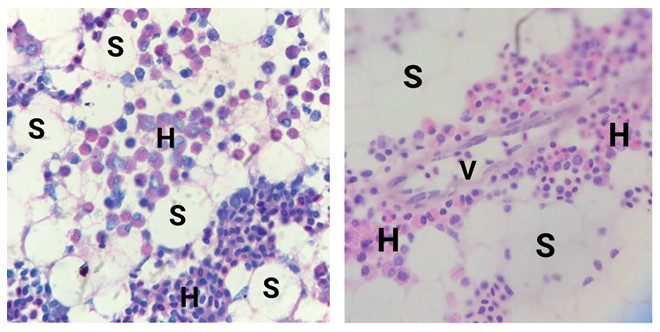
Результаты исследований и их обсуждение. В состав красного костного мозга входят три компонента: стромальный, сосудистый и гемопоэтический (рисунок 1). Стромальный компонент играет важную роль в опорной, трофической и регуляторной функции, создавая особое микроокружение для нормального развития кроветворных клеток. В его состав входят ретикулярные клетки, адипоциты, макрофаги [1].
Сосудистый компонент наряду с обычными сосудами микроцир-куляторного русла содержит синусоиды (венулярные синусы) - тонкостенные широкие 48 анастомозирующие сосуды, выстланные тонким эндотелием, через который в просвет синуса сквозь временно образующиеся в цитоплазме клеток поры поступают зрелые форменные элементы крови. Снаружи к синусоидам прилегают макрофаги и ретикулярные клетки.
Гемопоэтическая ткань костного мозга имеет вид шнуров, разделяется на дольки, состоящие из компактно упакованных клеток, образующих цилиндрические скопления вокруг артериол. Друг от друга дольки отделены дренирующими синусоидами. Ретикулиновые волокна и ретикулярные клетки составляют каркас костномозговых шнуров. Соприкасаясь друг с другом тонкими ветвящимися отростками, ретикулярные палетки образуют губчатую строму, в петлях которой расположены гемопоэтические клетки. Помимо ретикулярных клеток, фибробластов и кроветворных клеток в шнурах встречаются лаброци-ты (тучные клетки) и макрофаги, лежащие вблизи венозных синусов.
Рис. 1. Гистологическое строение костного мозга: А - окраска азур-2-эозин, (ув. х400), В - окраска гематоксилин-эозин (ув. х400), S - стромальный компонент, V - сосудистый компонет, Н - гемопоэтический компонент В костном мозге индеек выделяли 4 ростка гемопоэза: эритроцитарный, моноцитарный, гранулоцитарный и тромбоцитарный.
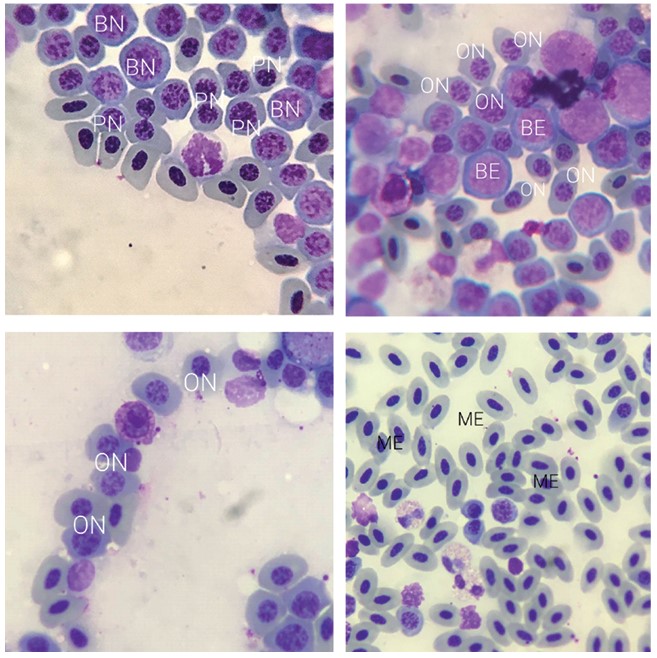
Эритроцитарный росток представлен созревающими эритроцитами на разных стадиях дифференцировки (рисунок 2). Первая стадия - базофильный эритробласт. Он имеет размер чуть больше нейтрофила, резко-базофильную цитоплазму и ядро, занимающее почти всю площадь. При окраске по Гимза ядро имеет фиолетовый цвет и рыхлый рисунок хроматина. Затем, при дифференцировке, цитоплазма и ядро клетки равномерно уменьшаются в размерах, достигая 6-9 мкм. Такая клетка называется полихроматофильным нормо-бластом. Она имеет ядро с более конденсированным рисунком, а её цитоплазма окрашена более нейтрально. Затем цитоплазма начинает увеличиваться в размерах и принимать овальную форму. Окраска цитоплазмы становится светло-коричневой, это связано с накоплением гемоглобина в ней. Ядро также принимает овальную форму, и в нем происходит конденсация хроматина. Клетка на данном этапе созревания называется ортохроматофильным нормобластом.
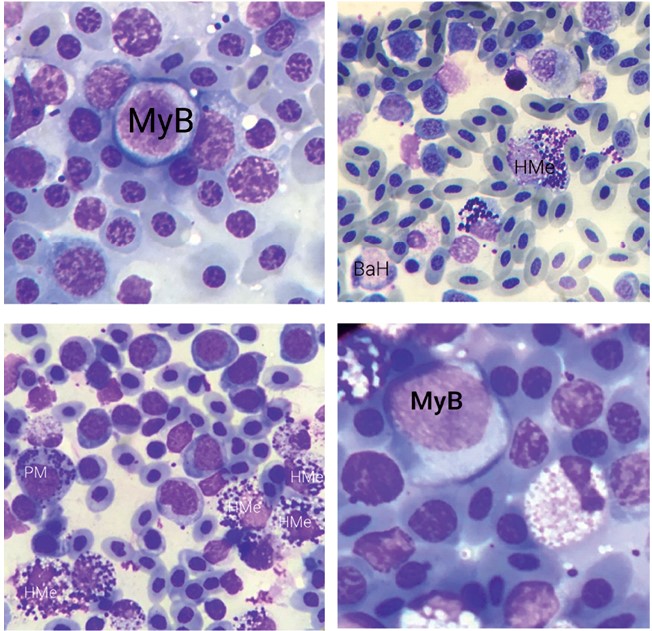
Рис. 2. Эритроцитарный росток (ув. х1000): BE - базофильный эритробласт, BN - базофильный нормобласт, PN - полихроматофильный нормобласт, ON - ортохроматофильный нормобласт, ME - зрелые эритроциты Лейкоцитарный росток представлен гранулоцитарной и моноцитарной линиями (рисунок 3) [7, 8]. Гранулоциты в процессе созревания разделяются на три ветви: базофильные, эозинофильные и псевдо-эозинофильные. Развитие гранулоцита начинается со стадии миело-бласта. Эта клетка имеет округлую форму цитоплазмы. Ядро также имеет правильную форму, занимает почти всю площадь цитоплазмы, рисунок хроматина в ядре - грубый, в ядре также визуализируется 2-3 ядрышка. Следующей стадией является стадия промиелоцита. Данная клетка имеет большую, чем миелобласт, площадь цитоплазмы, ядро чаще всего смещено к одному из полюсов, в цитоплазме виднеется первичная зернистость. Эти зерна имеют маленький размер, располагаются диффузно по всей площади клетки. Следующей стадией является миелоцит. На этом этапе клетка приобретает вторичную зернистость. Вторичные гранулы имеют характерную форму и окрас и позволяют дифференцировать клетку. В ядре хроматин становится более конденсированным, на данной стадии ядро может иметь более вытянутую форму. Следующей стадией является метамиелоцит. На этой стадии ядро имеет более вытянутую форму, в некоторых случаях - бобовидную. На следующих стадиях ядро сначала принимает палочковидную форму, а затем сегментируется. Данные стадии называются палочкоядерными, а затем сегментоядерными гранулоцитами.
Рис. 3. Миелопоэз (ув. х1000): MyB - миелобласт, PM - промиелоцит, HMe-гетерофильный метамиелоцит, BaH - палочкоядерный гетерофил Клетки моноцитарного ростка развиваются из монобластов. Эта клетка очень сильно внешне похожа на миелобласт, поэтому для их корректной дифференцировки используется электронный микроскоп [7]. Во время дифференцировки ядро принимает бобовидную форму, в цитоплазме появляются гранулы. После этого моноцит дифференцируется в клетки макрофагального ряда: гистиоциты, дендритные клетки.
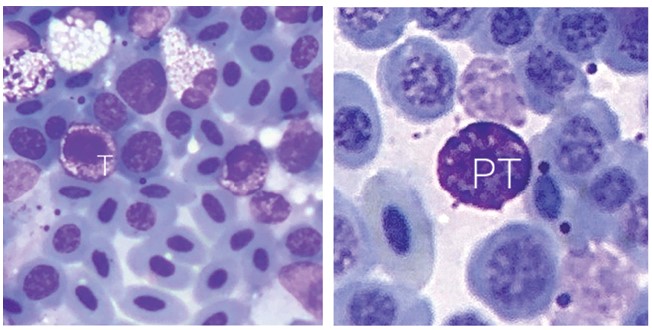
В отличие от млекопитающих, в костном мозге птиц отсутствуют мегакариоциты. Тромбоцитарный росток представлен тромбобласта-ми, протромбоцитами и тромбоцитами (рисунок 4). Эти клетки очень похожи между собой: при окраске по Романовскому-Гимза они имеют цитоплазму темно-фиолетового цвета с гранулами и ядро с плотно упакованным хроматином.
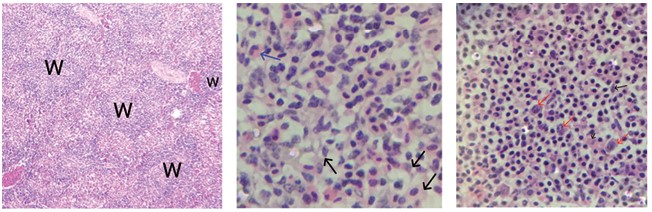
Рис. 4. Тромбопоэз (ув. х1000): PT - протромбоцит, Т - тромбоцит Селезенка птиц во время эмбрионального этапа развития выполняет гемопоэтическую функцию, однако в постнатальном онтогенезе данная функция уже утрачивается. Сформированная селезенка анатомически имеет бобовидную форму, немного сплюснутую снизу. На гистологическом препарате орган представлен красной и белой пульпой. Красная пульпа представлена большим количеством зрелых эритроцитов, её основная функция - депонирование крови. Белая пульпа представлена лимфоидными фолликулами (рисунок 5). В фолликулах происходит лимфопоэз, о чем свидетельствует наличие лимфобластов [2]. Основу стромального элемента составляют ретикулярные клетки. Они имеют зведчатую форму с большим количеством отростков, их основная функция - поддержание формы органа.
Рис. 5. Морфология селезенки: W - фолликулы белой пульпы; синими стрелками обозначены гранулоциты, черными стрелками - ретикулярные клетки, красными стрелками - лимфобласты Среди органов кроветворения и иммуногенеза особое внимание уделяется тимусу. Он является важнейшим органом иммунной защиты, являясь центральным звеном в системе иммуногенеза, и хорошо развит у молодых животных. Вилочковая железа у птиц располагается в области шеи, заключена в поверхностную фасцию, и залегает вдоль сосудисто-нервного пучка. Состоит из двух долей (правой и левой), каждая из которых разделена на гряды долек (тимические дольки) серовато-розового цвета. Количество долек зависит от возраста постнатального развития молодняка, породы и вида птицы, однако в среднем их 5-8 в каждой регионарной доле. У уток 3-4. Размер каждой тимусной дольки около 1 см в длину, форма эллипсоидная. Гистологически, ткани тимических долек дифференцированы на самостоятельные зоны: субкапсулярную, кортикальную, медуллярную и зону периваскулярных пространств. В медуллярной зоне у птиц, в отличие от рептилий, обнаруживаются тельца Гассаля, значение которых до настоящего времени остается неясным. Также как и у рептилий, у птиц тимус подвергается ранней возрастной инволюции, в последствии претерпевая тотальный лизис, и с периода полового созревания его лимфоидная паренхима замещается на жировую и соединительную ткани [6, 10].
В корковом и мозговом веществе тимуса неполовозрелой птицы возраста наблюдается количественное преобладание клеток более поздних стадий зрелости над менее дифференцированными элементами лимфоидного ряда. В корковом веществе преобладают малые формы тимоцитов. При этом, соотношение зрелых и незрелых форм остается в пороговом значении 1:5:11.
В мозговом веществе тимуса птиц, напротив, наиболее значительно по своему количеству малые тимоциты преобладают в двукратном объеме, в сравнении с земноводными и человеком. По данным исследований, митотическая активность клеток мозгового вещества снижена, в сравнении с кортикальным слоем. Данные по исследованиям приведены в таблицах 1 и 2.
Таблица 1 Количественное соотношение лимфоцитов разных стадий зрелости в тимусе птиц [6]
Как видно из приведенной таблицы, в кортикальном слое наблюдается увеличение числа больших тимоцитов, в сравнении с медуллярным слоем, что составляет 10,65% и 19,83% от общего числа клеток, соответственно. Количество средних тимоцитов медуллярного слоя на порядок ниже кортикального слоя, и составляет 27,65% и 22,8% от общего числа клеток, соответственно. Большое скопление малых тимоцитов наблюдалось в обеих исследуемых структурах. Процентные соотношения от общего числа клеток коркового и мозгового вещества идентичные, и составляют 66,55% и 66,50%, соответственно. Таблица 2 Морфометрические показатели коркового и мозгового вещества тимуса птицы
Клоакальная (Фабрициева) сумка представляет собой полостной мешкообразный лимфоидно-эпителиальный орган светло-серого цвета, связанный посредством короткого протока с клоакой. Данный орган первичного звена иммуннокомпетентной системы располагается в грудобрюшной полости под позвоночным столбом и имеет у кур 12-14 продольных складок, у голубей и уток - 2-4. Размер клоакальной бурсы вариабелен, в зависимости от вида и породы птицы и составляет примерно 1-2,5 см в диаметре. В каждой складке расположены один-два ряда лимфоидных ячеек (фолликулов), окруженных соединительнотканными элементами. В зависимости от функционального состояния в лимфоидной ячейке клоакальной сумки выделяют три зоны: кортикальную, пограничную и медуллярную. Также как и тимус, клоакальная сумка подвергается ранней возрастной регрессии и к началу морфофункциональной зрелости организма полностью исчезает [3, 5]. Аналог данного органа у млекопитающих отсутствует.
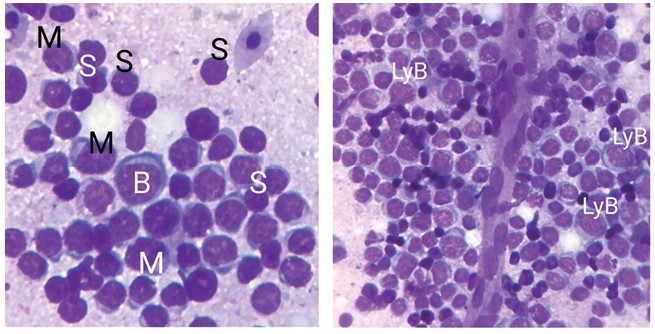
Данный орган характерен исключительно для птиц, его функцией является участие в созревании В-лимфоцитов (рисунок 6). Кроме вышеизложенного, в бурсе создается микросреда для расширения пула В-лимфоцитов и генерации разнообразного набора этих клеток, продуцирующих антитела [5]. Стенки органа состоят из слизистой, мышечной и серозной оболочек. Мышечная оболочка фабрициевой сумки является продолжением мышечной оболочки клоаки и образована двумя слоями мышечных пучков. Внутренняя оболочка сформирована циркулярным слоем гладкой мускулатуры, наружная - продольными пучками гладких миоцитов. Циркулярный слой более узкий, чем продольный, что характерно для уродеума - среднего отдела клоаки кур, в то время как в прямой кишке и в копродеуме клоаки наружный продольный слой уже, чем внутренний циркулярный. Слизистая оболочка образует продольные складки, различной длины и толщины, выступающие в просвет органа. Нами установлено наличие двух типов складок: высоких цилиндрических и низких конусовидных. Каждый лимфатический фолликул состоит из периферической корковой и центральной мозговой зон. Корковая зона заполнена малыми и средними лимфоцитами. Мозговая зона значительно светлее, и здесь находятся большие и средние лимфоциты. Тонкие механизмы созревания, пролиферации и антиген независимой дифференцировки В-клеток в бурсе Фабрициуса цыплят, экспрессии на их поверхности разнообразных генов, определяющих специфичность клеток, конверсия генов до сих пор окончательно не выяснены. Целенаправленные исследования дендритных секреторных клеток бурсы показали, что их предшественники появляются в сумке до попадания в бурсу B-клеток на 11-13 день эмбрионального развития и влияют на интенсивность развития зачатков фолликулов. При инволюции лимфоидные фолликулы бурсы теряют клетки сначала в корковой, а потом в мозговой зоне, происходит разрастание соединительнотканной стромы, появление кист, наблюдается сокращение плотности цитокератин-положительных ретикулоэпителиальных клеток медуллы (табл. 3). Таблица 3 Клеточный состав фабрициевой сумки птицы в постнатальном онтогенезе (%)
Рис. 6. Клеточный состав бурсы (ув. х1000): S - малые лимфоциты, M - средние лимфоциты, B - большие лимфоциты, LyB - лимфобласты Заключение. Проведенные исследования показывают, что у промышленной птицы основным органом гемопоэза и иммуногенеза является костный мозг, так как в нём созревают лейкоциты, эритроциты и тромбоциты. Селезёнка является основным органом лимфопоэза из-за наличия в ней лимфобластов. Красная пульпа селезёнки является депо красных кровяных телец. В фабрициевой сумке и тимусе происходит лимфопоэз и дифференцировка В- и Т- лимфоцитов, соответственно.
Список литературы:
Резюме. В статье представлены литературные и собственные данные о морфологическом строении, клеточном составе и функциях центральных органов иммунной системы птицы. В собственных исследованиях для изучения данных органов использовался гистологический и цитологический метод. Материалом для гистологического исследования служили кусочки органов, которые фиксировались в формалине, обезвоживались в спиртах, заливались в гистологический парафин и окрашивались гематоксилином-эозином, азуз-2-эозином. А для цитологического исследования материалом служили мазки-отпечатки с разрезов этих же органов, которые окрашивались по методике Май Грюнвальд-Гимза и монтировались под полистирол. В результате исследования было показано, что костный мозг является основным органом кроветворения. В нем происходит созревание эритроцитов, гранулоцитов, моноцитов и кровяных пластинок. Он состоит из стромального, сосудистого и гемопоэтического компонентов. Селезенка является одним из органов лимфопоэза, на что указывает наличие там большого количества лимфобластов. В красной пульпе селезенки также происходит депонирование эритроцитов, в то же время, в отличии от млекопитающих, селезенка не участвует в переработке старых эритроцитов. Тимус, или зобная железа, выполняет функцию дифференцировки Т-лимфоцитов. У птиц представлен цепочкой долек, лежащих вдоль трахеи. Фабрициева сумка (бурса) является органом, где происходит дифференцировка В-лимфоцитов. Она расположена над анусом, имеет белый цвет, овальную форму, на разрезе и гистологическом препарате представлена множественными длинными складками с лимфоидными образованиями. Также, в отличие от селезенки и костного мозга, тимус и бурса подвергаются редукции примерно на 130-150 дни жизни.
Ключевые слова: птица, иммунитет, костный мозг, тимус, селезенка, фабрициева сумка, гистология, цитология.
Сведения об авторах:
Михайлов Евгений Владимирович, кандидат ветеринарных наук, заведующий отделом экспериментальной фармакологии ФГБНУ «Всероссийский научно-исследовательский ветеринарный институт патологии, фармакологии и терапии»; 394087, г. Воронеж, ул. Ломоносова 114 б; тел.: 8-920-4025245; e-mail: voronezh81@rambler.ru.
Степанов Егор Максимович, аспирант, младший научный сотрудник лаборатории инновационных препаратов рекомбинантной протеомики ФГБНУ «Всероссийский научно-исследовательский ветеринарный институт патологии, фармакологии и терапии»; 394087, г. Воронеж, ул. Ломоносова 114 б; тел.: 8-950-7532764.
Ответственный за переписку с редакцией: Шабунин Борис Викторович, старший лаборант лаборатории инновационных препаратов рекомбинантной протеомики ФГБНУ «Всероссийский научно-исследовательский ветеринарный институт патологии, фармакологии и терапии»; 394087, г. Воронеж, ул. Ломоносова 114 б; тел.: 8-920-4167311; e-mail: bv.shabunin@gmail.com.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||


| 2011 © Ветеринария Кубани | Разработка сайта - Интернет-Имидж | |
|---|---|---|